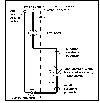
Functions
![]() Lat.
Zone
Lat.
Zone
|
Functional Examples of Cerebellar Pathways |
Now, let's figure out what cerebellar circuitry might accomplish. I will use the example of the visually guided movement of reaching and grasping an apple from an apple tree.
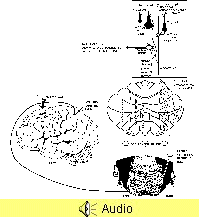
 The visual information
about the apple is carried from the retina to the cerebral
cortex via pathways that we will discuss later in the
course. This visual information eventually reaches a region
in the parietal lobe just caudal to the primary sensory
cortex called the posterior parietal cortex
(PPC; see illustration on next page). Cells in this cortical
area are involved in synthesizing the spatial coordinates of
contralateral body and space. For instance, there are cells
in the PPC that fire prior to moving the arm toward the
apple (desired object). These same cells would not
fire prior to the same arm movement that is not aimed at the
apple. Integration in the PPC leads to the programming of a
motor plan that will eventually result in the purposeful
movement of reaching out and grasping the apple. KEEP IN
MIND THAT NO REACH AND GRASP MOVEMENT HAS BEGUN-ONLY THE
PLANNING FOR SUCH A MOVEMENT.
The visual information
about the apple is carried from the retina to the cerebral
cortex via pathways that we will discuss later in the
course. This visual information eventually reaches a region
in the parietal lobe just caudal to the primary sensory
cortex called the posterior parietal cortex
(PPC; see illustration on next page). Cells in this cortical
area are involved in synthesizing the spatial coordinates of
contralateral body and space. For instance, there are cells
in the PPC that fire prior to moving the arm toward the
apple (desired object). These same cells would not
fire prior to the same arm movement that is not aimed at the
apple. Integration in the PPC leads to the programming of a
motor plan that will eventually result in the purposeful
movement of reaching out and grasping the apple. KEEP IN
MIND THAT NO REACH AND GRASP MOVEMENT HAS BEGUN-ONLY THE
PLANNING FOR SUCH A MOVEMENT.
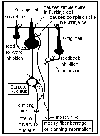
 Information from the
PPC is conveyed to the ipsilateral pontine grey
neurons via corticopontine fibers. Pontine grey neurons then
relay the planning information to the contralateral
lateral zone of the cerebellum via
pontocerebellar fibers, which everyone knows by now
are mossy fibers. Before the movement begins, cells in the
dentate nucleus are spontaneously firing, as are the mossy
fibers. The Purkinje cells are firing their simple spikes
(via the granule cells and their parallel fibers). Every
once in a while there is a climbing fiber complex spike in
the Purkinje cell. As the wave of "mossy fiber planning"
information hits the cerebellum, there will be an increase
in the firing of the dentate (due to the collaterals from
the mossy fibers) and an increase in the simple spikes fired
by Purkinje cells. The Purkinje cell message to the dentate
(increase in inhibition), and the dentate message to VA/VL,
will change due to this wave of mossy fiber planning
information. This new message headed for VA/VL contains
planning information for reaching out and grasping the
apple. The message in VA/VL is then relayed to the premotor
(PM) and supplementary motor areas (SMA), which in turn feed
into the primary motor cortex (area 4; the cells of origin
of the corticospinal tract). Remember all of this happens
before any movement has started!!
Information from the
PPC is conveyed to the ipsilateral pontine grey
neurons via corticopontine fibers. Pontine grey neurons then
relay the planning information to the contralateral
lateral zone of the cerebellum via
pontocerebellar fibers, which everyone knows by now
are mossy fibers. Before the movement begins, cells in the
dentate nucleus are spontaneously firing, as are the mossy
fibers. The Purkinje cells are firing their simple spikes
(via the granule cells and their parallel fibers). Every
once in a while there is a climbing fiber complex spike in
the Purkinje cell. As the wave of "mossy fiber planning"
information hits the cerebellum, there will be an increase
in the firing of the dentate (due to the collaterals from
the mossy fibers) and an increase in the simple spikes fired
by Purkinje cells. The Purkinje cell message to the dentate
(increase in inhibition), and the dentate message to VA/VL,
will change due to this wave of mossy fiber planning
information. This new message headed for VA/VL contains
planning information for reaching out and grasping the
apple. The message in VA/VL is then relayed to the premotor
(PM) and supplementary motor areas (SMA), which in turn feed
into the primary motor cortex (area 4; the cells of origin
of the corticospinal tract). Remember all of this happens
before any movement has started!!
 The dentate is
conveying PPC data that has gone through the computer
circuitry of the lateral cerebellum. This information
appears to be of importance in helping to start and stop
movements. Think of the lateral cerebellum and the dentate
as giving a boost to the corticospinal tract signal before
the actual signal starts. You all know that if you are
really "ready" to do a particular motor act, that you
usually do it faster and more accurately. Usually your
muscles contract fast and for shorter periods. Think of the
dentate as getting the corticospinal tract "ready" to reach
out and grab the apple (via the PM and SMA). Realize of
course, that when the information leaves the dentate, no
movement has started. Only planning information is destined
for the PM and SMA. This information is then relayed to the
primary motor cortex (MI) for the execution of movements via
the corticospinal tract.
The dentate is
conveying PPC data that has gone through the computer
circuitry of the lateral cerebellum. This information
appears to be of importance in helping to start and stop
movements. Think of the lateral cerebellum and the dentate
as giving a boost to the corticospinal tract signal before
the actual signal starts. You all know that if you are
really "ready" to do a particular motor act, that you
usually do it faster and more accurately. Usually your
muscles contract fast and for shorter periods. Think of the
dentate as getting the corticospinal tract "ready" to reach
out and grab the apple (via the PM and SMA). Realize of
course, that when the information leaves the dentate, no
movement has started. Only planning information is destined
for the PM and SMA. This information is then relayed to the
primary motor cortex (MI) for the execution of movements via
the corticospinal tract.
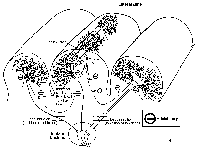 You might wonder what
all of those crazy inhibitory cells in the cerebellar cortex
of the lateral zone are doing. Well, the feedback
inhibition from the Golgi cell (see A above) could be a
temporal (timing) control for sharpening the initial
excitatory mossy barrage. That is, maybe for the cerebellum
to do its "thing", only the very first part of the mossy
fiber barrage of planning data is let through to the
Purkinje cells. The feedforward inhibition via the
basket cells (see B above) is involved in spatially
sharpening the planning of information signals. That is, as
the parallel fibers associated with the mossy fiber barrage
excite the Purkinje cells, the basket cells are inhibiting
Purkinje cells on the flanks of the active zone. This would
kind of emphasize a "hot" zone of Purkinje cell activity.
The different firing rates and patterns of dentate neurons
driven by the "hot" zone and the flanking zones are then
conveyed to VA/VL, the PM and SMA, and in turn to MI (the
cells of origin of the corticospinal tract). The cerebellum
uses this spatial information in the planning of movements
headed for the cells of origin of the corticospinal tract.
You might wonder what
all of those crazy inhibitory cells in the cerebellar cortex
of the lateral zone are doing. Well, the feedback
inhibition from the Golgi cell (see A above) could be a
temporal (timing) control for sharpening the initial
excitatory mossy barrage. That is, maybe for the cerebellum
to do its "thing", only the very first part of the mossy
fiber barrage of planning data is let through to the
Purkinje cells. The feedforward inhibition via the
basket cells (see B above) is involved in spatially
sharpening the planning of information signals. That is, as
the parallel fibers associated with the mossy fiber barrage
excite the Purkinje cells, the basket cells are inhibiting
Purkinje cells on the flanks of the active zone. This would
kind of emphasize a "hot" zone of Purkinje cell activity.
The different firing rates and patterns of dentate neurons
driven by the "hot" zone and the flanking zones are then
conveyed to VA/VL, the PM and SMA, and in turn to MI (the
cells of origin of the corticospinal tract). The cerebellum
uses this spatial information in the planning of movements
headed for the cells of origin of the corticospinal tract.
Lesions of the lateral zone of the cerebellum and dentate result in errors in the direction, force, speed, and amplitude of movements. (Interestingly, these incoordinated movements are being carried out by a healthy corticospinal tract. It is just that the corticospinal tract is receiving bad information). The "priming" of the corticospinal tract by the dentate is lost. It takes longer to get the movement going, the appropriate muscles contract for too long, and the movement stops too late. There is a deterioration of coordinated movements called decomposition of movement. Instead of a nice smooth movement, the movement consists of jerky parts of the movement. Dysmetria, also called past-pointing, occurs when the patient tries to touch their nose with their finger. Rapid alternating movements (pronation/supination) is awkward and is called dysdiadochokinesis and there is the rebound phenomena. Hold the patients tensed arm away from his/her sternum and chest and then let go. It will fly into their sternum. There also is tremor during the incoordinated movement. This is called movement, or intention tremor. It is hard for me to explain why this might occur with lateral zone lesions but it might be due to the fact that there are problems in correctly stopping movements. This results in the movement going too far, after which there is a correction and then oscillations occur.